
AUTHOR:
Allen MacNeillSOURCE: Original essay
COMMENTARY: That's up to you...
John Wilkins over at
Evolving Thoughts has posted the following abstract, along with a brief discussion of its significance for evolutionary biology:
The Nature of Plant Species
Loren H. Rieseberg [1], Troy E. Wood [1] and Eric J. Baack [1]
Many botanists doubt the existence of plant species, viewing them as arbitrary constructs of the human mind, as opposed to discrete, objective entities that represent reproductively independent lineages or 'units of evolution'. However, the discreteness of plant species and their correspondence with reproductive communities have not been tested quantitatively, allowing zoologists to argue that botanists have been overly influenced by a few 'botanical horror stories', such as dandelions, blackberries and oaks. Here we analyse phenetic and/or crossing relationships in over 400 genera of plants and animals. We show that although discrete phenotypic clusters exist in most genera (> 80%), the correspondence of taxonomic species to these clusters is poor (< 60%) and no different between plants and animals. Lack of congruence is caused by polyploidy, asexual reproduction and over-differentiation by taxonomists, but not by contemporary hybridization. Nonetheless, crossability data indicate that 70% of taxonomic species and 75% of phenotypic clusters in plants correspond to reproductively independent lineages (as measured by postmating isolation), and thus represent biologically real entities. Contrary to conventional wisdom, plant species are more likely than animal species to represent reproductively independent lineages.
Nature 440, 524-527 (23 March 2006) | doi:10.1038/nature04402; Received 26 July 2005
[1] Department of Biology, Indiana University, Bloomington, Indiana 47405, USA. Correspondence and requests for materials should be addressed to: Loren H. Rieseberg (Email: lriesebe@indiana.edu).
He pointed out that this would mean that botanists would seem to have a clearer definition of what a species is than most zoologists. However, the paper cited above notwithstanding, I think I would have to disagree with that assertion.
For over a century, evolutionary biologists have been wrestling with the concepts of species and
speciation, trying to determine which (if any) of the various species concepts are most useful, and which mechanism of speciation is the most important in nature. But all of this has been predicated on the assumption that species actually exist outside of the human imagination. Is it possible that all of this has been an exercise in futility, a kind of "
origin of the specious?"
Consider the fact that nearly all of the people whose opinions on the subject of species and speciation have themselves studied animals almost exclusively, especially Charles Darwin, Theodosious Dobzhansky, and Ernst Mayr. Not really surprising: after all, we're all animals and most interested in ourselves. Furthermore, the most widely applied definition of a species, the so-called "
biological species concept" was most forcefully advocated by Ernst Mayr in
Systematics and the Origin of Species.
Which leads to the following question:
• Is there something peculiar about animals that predisposes people who study them to frame the question of what a species is in such a way as to get the biological species concept as the answer?
In a word, yes:
Lynn Margulis at the University of Massachusetts has argued forcefully for the following radical interpretation of the "origin of species":
• There are no such thing as "species" among the most numerous and diverse organisms on Earth: the prokaryotes (commonly referred to as "bacteria") either don't have species at all in any commonly accepted sense, or they are all one huge "species."
• Horizontal gene transfer (i.e. gene transfer from organism to organism without sex, reproduction, and therefore "descent") hopelessly muddies the phylogenies of whole kingdoms of organisms (including many animals, and possibly including ourselves).
• In particular, the Protoctists (i.e. unicellular eukaryotes, also referred to as Protists) have such diverse and bizarre sexual and reproductive behaviors as to be all but unclassifiable (some have as many as eight "sexes"!)
• Nearly all fungi and many plants are generally indifferent to species boundaries. As sessile organisms, fungi and plants they can't be choosy about whom to mate with; whomever is nearby will do.
• This is particularly true for the fungi, who very rarely "go outside" - they live nearly their entire lives underground as widely distributed networks of tubular cells called hyphae.
• Orchids have been known to hybridize across not only species lines, but across genera and even families.
• Nearly all plants have the ability to reproduce asexually; indeed, some (like the dandelion) have given up sexual reproduction entirely and are therefore in essentially the same category as bacteria.
Pretty radical stuff, and a kind of "universal acid" for the
biological species conceptfor anything except animals (but watch out for
whiptail lizards).
Lynn Margulisis admittedly a radical in this regard, but one with a solid track record. Her theory for the evolution of eukaryotes, called "
serial endosymbiosis", was considered by many to be the ravings of a lunatic when she first proposed it in 1969. Now it is the most mainstream of orthodoxies.
So, how about it: are non-animal species "real"? Consider the following (From: Margulis, Lynn and Sagan, Dorion (2002).
Acquiring Genomes: A Theory of the Origin of Species, Basic Books, New York, NY, pp. 54-55):
For the numbers of living species…we have only crude estimates that may be wildly incorrect. Animals, probably because people are good at distinguishing beetles, dominate. Over 10 million - perhaps as many as 30 million - are thought to exist. Some 500,000 plants, 100,000 fungi, and 250,000 protoctists are suggested to be lurking in the woods and waters of this world."
As for bacteria, although thousands have been named as species and no doubt thousands can be distinguished, the [biological] species concept doesn't apply. Although bacteria can be grouped on the basis of common features, these groups change so quickly that they are never fixed and recognizable like eukaryote species. Bacteria pass genes back and forth. All can simply reproduce, and thus at any given time have but a single parent. The intervention of sex…is a unidirectional affair. The genes pass from a donor to a recipient…but donors can change to recipients and vice versa in minutes. Furthermore the gene swapping is entirely optional…[I]ndeed bacteria are willing and able to "have sex" with naked DNA molecules that they absorb from the water in which they are bathed.
Life originated with bacteria; therefore we can say that the origin of life was concurrent with the origin of bacteria. But we agree with Professor Sorin Sonea and his colleague Lucien Mathieu, of the Université de Montreal, that bacteria do not have species at all (or, which amounts to the same thing, all of them together constitute one single cosmopolitan species). Speciation is a property only of nucleated organisms.
The Taxonomy of Some Common TreesOkay, so perhaps taxonomists have been missing the forest for looking too closely at the trees? Consider these seemingly obvious tree species (if you live in the northeastern United States, you can see most of them by simply stepping outside and looking around):
SPECIES:
Acer saccharum
COMMON NAMES :
• sugar maple
• rock maple
• hard maple
TAXONOMY :
The currently accepted scientific name of sugar maple is
Acer saccharum Marsh. Sugar maple is highly variable genetically and taxonomic controversy abounds. Some taxonomists recognize two to six varieties, but others recognize these entities as forms or subspecies. Several ecotypes or races, each exhibiting clinal variation, have also been delineated.
Florida maple (
A. barbatum), chalk maple (
A. leucoderme), and black maple (
A. nigrum) hybridize and intergrade with sugar maple and are often included in the sugar maple complex. Some authorities recognize these taxa as subspecies of sugar maple, but most delineate them as discrete species. Sugar maple hybridizes with red maple (
A. rubrum) in the field, and with bigleaf maple (
A. macrophyllum) under laboratory conditions.
Acer X senecaense Slavin is a hybrid derived from an
A. leucoderme x sugar maple cross.
A. skutchii is closely related to sugar maple and is treated as a subspecies by some taxonomists.
AUTHORSHIP AND CITATION :
Tirmenstein, D. A. 1991.
Acer saccharum In: U.S. Department of Agriculture(2002, September), Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory Fire Effects Information System:
http://www.fs.fed.us/database/feis/plants/tree/acesac/introductory.html
SPECIES:
Quercus alba
COMMON NAMES :
• white oak
• stave oak
• ridge white oak
• forked-leaf white oak
• fork-leaf oak
TAXONOMY :
The currently accepted scientific name of white oak is
Quercus alba L. It is a member of the order Fagales and has been placed within the white oak subgenus (Lepidobalanus). Three varieties of white oak are commonly recognized:
•
Quercus alba var.
alba•
Quercus alba var.
repanda Michx.
•
Quercus alba var.
latiloba Sarg.
Some authorities recognize these entities as forms rather than varieties.
White oak is highly variable genetically, and many forms and ecotypes have been described. According to Fowells, "no definite races have been defined, but within such a tremendously diverse habitat, climatic races undoubtedly exist." White oak readily hybridizes with many other species within the genus
Quercus, including swamp white oak (
Q. bicolor), bur oak (
Q. macrocarpa), chinkapin oak (
Q. muehlenbergi), dwarf chinkapin oak (
Q. prinoides), overcup oak (
Q. lyrata), swamp chestnut oak (
Q. michauxii), sandpost oak (
Q. margaretta), chestnut oak (
Q. prinus), English oak (
Q. robur), Durand oak (
Q. durandii), and post oak (
Q. stellata). Hybrids, their common names, and purported origins are listed below.
• Beadle oak……X beadlei Trel. (
Quercus alba x
michauxii)
• Bebb oak……..X bebbiana (
Q. alba x
Q. macrocarpa)
• Deam oak…….X deamii (
Q. alba x
Q. muehlenbergi)
• Faxon oak……X faxonii Trel. (
Q. alba x
Q. prinoides)
• Fernow oak…..X fernowii Trel. (
Q. alba x
Q. stellata)
• Jack oak……...X jackiana Schneid. (
Q. alba x
Q. montana)
• Saul oak……...X saulii Schneid. (
Q. alba x
Q. prinus)
Saul oak was formerly known as
Q. alba f.
ryderii but is now considered a heterozygous hybrid form of white oak.
Introgressive populations are locally common throughout much of the range of white oak. Hybrid swarms derived from complex mixtures of parental forms are particularly common on disturbed sites, at the margins of white oak's range, and where several oak species occur sympatrically.
AUTHORSHIP AND CITATION :
Tirmenstein, D. A. 1991. Quercus alba In: U.S. Department of Agriculture(2002, September), Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory Fire Effects Information System:
http://www.fs.fed.us/database/feis/plants/tree/quealb/introductory.html
SPECIES:
Salix nigra
COMMON NAMES :
• black willow
• swamp willow
• southwestern black willow
• Gulf black willow
• scythe-leaved willow
TAXONOMY :
The currently accepted scientific name of black willow is
Salix nigra Marsh. Recognized varieties are
S. nigra var.
nigra Marsh.,
S. nigra var.
altissima Sarg.,
S. nigra var.
falcata (Pursh.) Torr., and
S. nigra var.
lindheimeri.
Salix nigra,
S. gooddingii Ball, and
S. amygdaloides Anderss. are closely related taxa commonly referred to as the black willows. The three species are not easily distinguished morphologically, and in fact, some authorities consider
S. gooddingii to be
S. nigra var.
vallicola Dudley or
S. n. var.
venulosa (Anderss.) Bebb.
S. amygdaloides is sometimes considered to be
S. nigra var.
amygdaloides Anderss. For our purposes, however, these varieties will be considered as separate species.
S. nigra hybridizes with
S. amygdaloides (
S. X
glatfelteri Schneider);
S. alba (
S. X
hankensonii Dode); and
S. lucida (
S. X
schneider Boivin).
AUTHORSHIP AND CITATION :
Tesky, Julie L. 1992. Salix nigra In: U.S. Department of Agriculture(2002, September), Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory Fire Effects Information System:
http://www.fs.fed.us/database/feis/plants/tree/salnig/introductory.html
The Taxonomy of RosesAh, but according to Gertrude Stein, "a rose is a rose is a rose", right? Well, check these out:
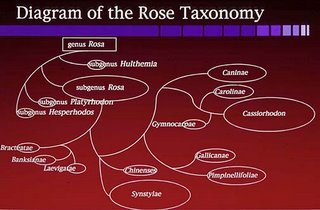
Relationships Among Some Species of Roses
(From: Ma, Y., Crane, C. F., and Byrne, D.H. (1997). Relationships among some Rosa species, Caryologia, vol. 50, n. 3-4, pp. 317-326):
The genus Rosa is widely distributed and taxonomically difficult. The great majority of its 200 species hall into the ten sections of subgenus Rosa; the three other subgenera contain one to three species each. Morphological and ecological variation with subgenus Rosa is almost continuous because of wide adaptation, wide limits of crossability and hybrid fertility, and frequent human intervention in bybridization and dispersal. Polyploidy is frequent in sections Pimpinellifoliae, Gallicanae, Cinnamoneae, and Caninae of subgenus Rosa and essentially absent elsewhere among the wild species. Karyotype analysis is a traditional first step in the comparison of genomes among related species that are to be included in a breeding program.
Think that's bad? Consider this "war of the roses":
Chromosomes in Relation to Sterility in Roses
(From: Pal, B. P, (1972). The Rose in India, 2nd Ed. Maggs Bros, Ltd, New Delhi, RI)
The basic number of chromosomes in roses is 7, and several important species of Asian origin, which have contributed significantly to the development of such famous groups as the Hybrid Perpetual and the Hybrid Teas, are diploids, having 14 chromosomes (7 maternal and 7 paternal). These include moschata, gigantea, multiflora, wichuraiana and chinensis. A number of Western species with which these Asian diploids crossed to yield several modern groups of roses are tetraploids, with 28 chromosomes. These, which had their chromosome number doubled during the course of their evolution, whose record we do not have, include gallica, foetida and their derivatives such as damascena and centifolia.

One of the more important crosses involving these species of diverse geographical distribution was that between gallica and a variety combining in it the genes of chinensis and gigantea. The hybrid arising directly from this cross was a triploid, having 21 chromosomes, 14 from the European parent and 7 from the Asian. This imbalance made it sterile and restricted its utility; but plants with 28 chromosomes arose spontaneously from it. They were fertile and gave rise to the Hybrid Perpetuals with the further incorporation of some damascena genes.

A triploid origin has also been shown for the Hybrid Teas, which followed and replaced to a large extent the Hybrid Perpetual. The Hybrid Teas were derived from a cross between the tetraploid Hybrid Perpetuals and a diploid variety of the Tea roses which combines in it the genes of three Asian (moschata, chinensis and gigantea) and one European (damascena) species. As in the case of the Perpetuals, tetraploid varieties were spontaneously obtained from the original triploid hybrid; this restored the fertility of the plant, making it possible to obtain many different combinations of genes in its progeny.

The condition of triploidy resulting from the crossing of diploid and tetraploid parents, associated as it was with hybrid sterility, delayed the release of gene combinations which were destined to develop into our modern groups of roses. Not all triploid hybrids, however, had to wait for a change to the tetraploid level before their usefulness could be appreciated…There are also examples where a cross involving a diploid and a tetraploid skipped the expected condition of triploidy and gave rise to the tetraploid state directly through an abnormal behavior of the diploid parent, which passes its entire complement of chromosomes to its sex cells in place of only half the number, which is the normal practice.
Ouch! Those roses don't seem to give a damn about chromosome number, polyploidy, and all of those other things that are so often developmentally fatal to most animals. But, you say, this mess is mostly the result of humans messing about with the chromosomes of the original rose. Let's look at some roses that haven't been so corrupted by bourgeois gardeners. Say, some good, proletarian Russian roses...
The Present State of Taxonomy of the East European Roses
(From: Schanzer, L.A. (2001) Biological Series, vol. 106, part 2, pp. 1-2)
http://herba.msu.ru/russian/journals/bmsn/106/2001_2.html
Critical overview of the taxonomic literature on the East European species of Rosa leads to the conclusion that this genus is still extremely inadequately studied as to species composition and relations. Quite a number of species, infra-, and superspecific taxa described so far do not make the taxonomy and nomenclature of the group any more clear. They moreover make any firm determination of most of the species perfectly impossible. Data available on chromosome numbers, interspecific crossings, and compatibility of West European species point to the necessity of such studies of East European species as well. The latter remain completely unstudied in these respects so far. On the other hand, sparse data on infrapopulational variability of Rosa suggest such studies to be important to understanding of many disputable questions of the Rosa taxonomy."
Conclusion: A rose by any other name would smell as sweet, but that doesn't help us with the classification of roses.
Perhaps Darwin's most important insight was his realization that species are not immutable, that they can intergrade over time in an "insensible series." But what Darwin didn't have the courage to come right out and say, and what most evolutionary biologists in general don't have the courage to propose, is that there are really no such thing as species at all, at least not in the way we have traditionally defined them. Darwin should have realized this: he made it clear that natural selection happens at the level of individuals, never at the level of species. Evolutionary biologists have agreed with him, but have not taken the obvious next step: to declare that individuals living organisms are the only things that exist in the natural world, and that species (including animal species) may quite literally be figments of the human imagination.
Labels: allopatric, biological species concept, ecological species concept, geographic isolation, morphological species concept, phylogenetic species concept, speciation, species, sympatric

















