The Searchers

AUTHORS: William A. Dembski and Robert J. Marks II
SOURCE: Proceedings of the 2009 IEEE International Conference on Systems, Man, and Cybernetics. San Antonio, TX, USA – October 2009, pp. 2647-2652
COMMENTARY: Allen MacNeill
First, congratulations to Drs. Dembski & Marks! Publication is the life blood of all career academics and the living heart of the intellectual process. It takes courage and hard work (and a little bit of luck) to get your original work published, and more of the same to weather the criticism that inevitably ensues. But, just as one cannot have a fencing match without an opponent, real progress in any intellectual endeavor cannot come from consensus, but only from the clash of ideas and evidence.
And so, to specifics:
I have no quibble with most of the mathematical analysis presented. Indeed, given the assumptions upon which the authors' Conservation of Information (COI) theory is based (with which I do not necessarily agree, but which are clearly presented in their paper), the analysis presented is apparently not completely outside the domain of No Free Lunch (NFL) theorems in general.
However, the same cannot be said for the application of these ideas to biological evolution. To be specific, consider the following quote [Dembski & Marks (2009) pg. 2651, lines 2-5]:
"From the perspective of COI, these limited number of endpoints on which evolution converges constitute intrinsic targets, crafted in part by initial conditions and the environment." [emphasis added]
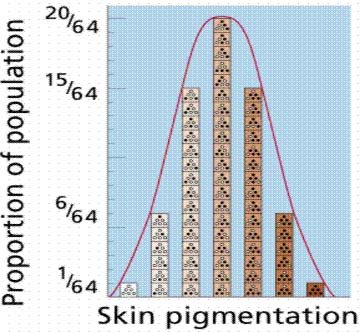
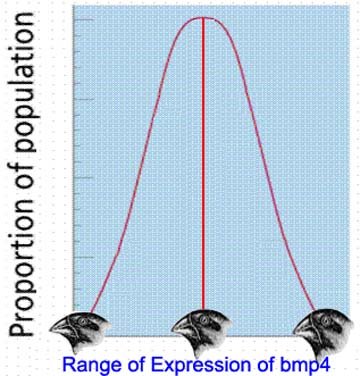
This is indeed the crux of the issue vis-a-vis biological evolution. While it is clearly the case that Simon Conway-Morris asserts that there is an apparently limited number of biological "endpoints", it is neither the case that Morris' viewpoint represents the core of evolutionary theory, nor that his point is relevant to the analysis of COI presented in Dr. Dembski and Marks' paper.
To be specific, the highlighted qualifier from the quote above – crafted in part by initial conditions and the environment – is precisely the issue under debate between evolutionary biologists and supports of intelligent design (ID).
Taken at face value, this qualifying simply phrase means that, given specific starting conditions and a specific time-varying environmental context, the various mechanisms of evolution (e.g. mutation, natural selection, genetic drift, inbreeding, etc.) tend to converge on a relatively limited set of genotypic and phenotypic "endstates" (i.e. what could be loosely referred to as "evolutionary adaptations").
This is simply another way of defining evolutionary convergence, and in no way constitutes evidence for intrinsic evolutionary teleology. On the contrary, it simply provides support for the hypothesis that, given similar conditions, similar outcomes result.
Furthermore, it assumes that virtually all characteristics of living organisms are adaptations (that is, genotypic/phenotypic characteristics that fulfill some necessary function in the lives of organisms). However, this is manifestly not the case, nor is it an absolutely necessary component of current evolutionary theory. On the contrary, many (perhaps the majority) of the characteristics of living organisms are not adaptive. This is certainly the case at the level of the genome, as evidenced by the neutral and nearly neutraltheories of molecular evolution.
Finally, Morris' (and, by extension, Dembski and Marks') position completely omits any role for historical contingency, which both the fossil and genomic record indicate are of extraordinary importance in macroevolution. As Dembski and Marks state, the "endpoints" (perhaps it would be more precise to refer to them as "way stations") of macroevolution depend fundamentally on initial conditions and the environment. But this is not fundamentally different from Darwin's position in the Origin of Species:
"The complex and little known laws governing variation are the same, as far as we can see, with the laws which have governed the production of so-called specific forms. In both cases physical conditions seem to have produced but little direct effect; yet when varieties enter any zone, they occasionally assume some of the characters of the species proper to that zone." [Darwin, C. (1859) Origin of Species, pg. 472, emphasis added]
Moreover, Dembski and Marks' analysis completely ignores the appearance (or non-appearance) of new genotypic and phenotypic variations, and on the accidental disappearance of such characteristics (via extinction), without regard to the adaptive value of such characteristics, or the lack thereof.
In other words, Dembski and Marks' analysis, while interesting from the standpoint of what could be called "abstract" search algorithms, completely fails to address the central issues of evolutionary biology: the source of evolutionary novelty (i.e. the "engines of variation"), the effects of changing environmental conditions on the actual forms and functions of living organisms, and the fundamental importance of historical contingency in the ongoing evolution of genotypes and phenotypes.
************************************************
As always, comments, criticisms, and suggestions are warmly welcomed!
--Allen
Labels: conservation of information, engines of variation, evolution, intelligent design, macroevolution, no free lunch theorems, Robert Marks, search algorithms, William Dembski
posted by Allen MacNeill @ 12/07/2009 08:51:00 PM
6 comments
![]()
![]()